¿QUE ES LA VIDA? Erwin Schrödinger, 60 años después
Erwin Schrödinger en hace una pregunta ¿por qué los átomos son tan pequeños?, a lo que comenta, que los procesos físicos y químicos básicos para el funcionamiento de un organismo requieren la participación de grandes cantidades de átomos para realzarse de modo seguro y es por esto que una estructura con pocos átomos no podría permitir procesos biológicos elementales.
La sorpresa es que un organismo vivo se mantiene ordenado a pesar de la agitación térmica y otros comportamientos que tienden a desordenar cualquier estructura constituida por muchas partículas, entonces como es que se puede crear y mantener el orden de un ser vivo?, Schrödinger explica que existen dos formas, el orden a partir del desorden y orden a partir del orden, donde la primera implica que un organismo se ordena por una pérdida del orden del ambiente, la segunda que el orden se transmite genéticamente por el ADN. Esto lo explica Max Delbrück, diciendo que moléculas grandes formadas por uniones covalentes tienen suficiente estabilidad como para almacenar información genética, opina que la vida es un proceso diferente único que será explicado por las leyes físicas. En conclusión la biología será útil a la física permitiendo descubrir nuevas leyes, y también la física a la biología ofreciendo una explicación unificada de la vida
LOS PRIMEROS SISTEMAS VIVOS, UNA PESTECTIVA BIOENERGETICA.
Se han encontrado meteoritos provenientes de la corteza marciana con inclusiones de carbonato, siendo interpretado como un origen bioenergético, haciendo pensar que probablemente la materia orgánica haya sido traída a la superficie terrestre desde un meteorito.
Los hidrocarburos y sus derivados estuvieron presentes en la reserva orgánica de la tierra primitiva, permitiendo la formación de membranas por organismos primitivos. Parece ser que los sistemas vivos se desarrollaron en soluciones diluidas de algunos sustratos como aminoácidos, carbohidratos, y bases puricas. Una forma de captura de energia es por reacciones de oxido reducción, teniendo a gradientes iónicos como fuentes potenciales de energía
TERMODINAMICA Y CALORIMETRIA DE LOS SISTEMAS VIVOS.
Calorimetría de los sistemas vivos y la termodinámica clásica desarrollada en paralelo, a partir de experimentos de hielo a principios de Lavoisier calorímetro en conejillos de indias, seguida por la investigación macrocalorimetria Dubrunfaut de los procesos de fermentación y Atwater-Rosa, toda la calorimetría cuerpo en humanos y animales domésticos, a la introducción de la famosa Tian instrumento Calvet encontró que la entrada en los campos tan diversos de la biología.
En este trabajo, se presentan seis ejemplos de la vida-calorimetría sistema y la termodinámica. Estos son: (i) oscilaciones glucolítica lejos del equilibrio termodinámico, (ii) el crecimiento y los balances de energía en la fermentación y la respiración cultivos de levaduras, (iii) el control calorimétrico directos e indirectos del metabolismo de los reptiles estimuladas eléctricamente, (iv) los factores climáticos y biológicos que influyen en la constancia de la temperatura y la distribución en el montículo de una colonia de hormigas de madera como un ejemplo de un sistema ecológico complejo, (v) las consideraciones energéticas en la agrupación de abejas europeas en el invierno como un medio para ahorrar energía y los alimentos almacenados, así como por su japonés homólogos en la defensa contra los depredadores Hornet, y (vi) los aspectos energéticos y la evolución de la tasa de la masa específica de producción de entropía, la disipación de los llamados de la envolvente o psiu función.
Los ejemplos presentados son sólo una selección muy personal de los sistemas vivos de un amplio espectro en todos los niveles de complejidad. Común para todos ellos es que fueron investigados calorimétricamente en el fondo de la termodinámica clásica e irreversible
EL OLFATO Y SUS RECEPTORES. LA HISTORIA DE UN NOBEL.
El olfato es considerado como el órgano mas desarrollado al momento del nacimiento. Pero la historia de este sentido empieza mucho tiempo atrás, hace casi 3.500 millones de años, durante la aparición de la vida en la tierra, las primeras células desarrollaron un sentido químico para percibir información que les llegaba de su entorno, en olfato es el sentido mas primitivo y el primero en aparecer en la escala evolutiva, tras evolucionar en los peces, este es muy importante en todos los organismos, en el hombre, el olfato sufre una serie de aprendizaje hasta la edad de 20 años, manteniéndose estable hasta los 40 años y empezando a decaer a partir de los 50, numerosos estudios han demostrado que en la mujer el olfato esta mas desarrollado y al envejecer lo pierde en menor grado que los hombres. Es además, un sentido con numerosas interconexiones con los centros de la memoria y de las emociones.
Las principales causas de la alteración o perdida del olfato, ya sea temporal o permanente, son resfriado común, inflamación de la mucosa nasosinunal, los traumatismos craneofaciales, el tabaquismo y las enfermedades neurodegenerativas. Las segundas causas menos frecuentes son las genéticas, los medicamentos, la cocaína, la exposición a tóxicos y contaminantes o los factores nutricionales. Los resultados preliminares de un estudio epidemiológico reciente (OLFACAT, 2003), realizado en una amplia muestra de población general catalana, demuestra que el 1% de la población presenta una perdida total del olfato (anosmia), mientras casi un 20% presenta una perdida parcial (hiposmia).
A partir de la publicación del la estructura del ADN, los trabajos en el área genética empezaron a hacerse mas importantes en la en el transcurso de la investigación científica, antes de los años 90, ya se conocía la localización de la mucosa olfatoria, sus pigmentos e incluso las células que lo formaban, en los años 80, se realizan estudios importantes sobre este sentido: se obtuvo y almaceno ADN de mucosa olfatoria y bibliotecas de genes y se demostró que al estimular los cultivos de las células olfatorias aumentaba la concentración de cAMP, un segundo mensajero que mediaba la estimulación de receptores adrenérgicos y colinérgicos ligados a las proteínas G.
En las ultimas décadas los trabajos realizados sobre el olfato fueron números, durante los años 90, Linda Buck y Richard Axel, continuaron con los estudios sobre este sentido, cada uno por separado, los mecanismos del olfato estaban regulados por una superfamilia de mas de 1.000 genes y las neuronas expresaban solo un tipo de receptor olfativo, estableciendo la teoría “de la llave y el candado”, otros estudios también han tenido gran importancia en esta rama, no obstante en el año 2004 se otorgo un reconocimiento a los pioneros en el descubrimiento de la superfamilia de los genes que codifican los receptores olfatorios, no obstante, hoy en día, aun existen muchas preguntas relacionadas con el olfato, que se esperan responder con mas estudios de este sentido muy complicado, el olfato.
viernes, 2 de abril de 2010
miércoles, 31 de marzo de 2010
CUESTIONARIO
1.- ¿Qué es la bioenergética?
Es el análisis cuantitativo de la forma en que los organismos adquieren y utilizan la energía, la bioenergética puede considerase una parte especial de la ciencia de las transformaciones energéticas denominada termodinámica.
2.- ¿Qué es un sistema termodinámico?
Un sistema termodinámico, es la parte del universo que elegimos para el estudio, tal como un recipiente de reacción, un organismo, una única célula bacteriana, el resto del universo se considera como el entorno, este debe tener unos límites definidos.
3.- ¿Por qué el ATP es muy importante en los sistemas vivos?
Las células requieren un continuo suministro de energía. Esta es necesaria para la síntesis de moléculas complejas, la ejecución de trabajo mecánico y el transporte de sustancias a través de sus membranas. La energía es transferida desde las reacciones químicas que la acumulan a las que las consumen, mediante una molécula especial, el ATP. En las células, la energía que recibe o cede el ATP es la contenida en el enlace entre su último fosfato y el resto de la molécula. El enlace se forma durante la síntesis de ATP: incorporan así energía, la que se cede cuando el enlace se escinde.
4.- ¿Mencione la relación que hay entre el ATP y la energía libre?
En la hidrólisis del fosfoenolpiruvato, el bifosfoglicerato , el adenosina trifosfato y el pirofosfato, los dos productos de la hidrólisis llevan una carga negativa.
La repulsión entre estos productos iónicos favorece intensamente la reacción de hidrólisis
La hidrólisis se ve favorecida a veces no sólo por la estabilización de la resonancia del ortofosfato sino también por la capacidad del otro producto para adoptar más formas moleculares, lo cual contribuye a la hidrólisis del del fosfoenolpiruvato ya que el piruvato puede tautomerizarse, con la liberación de energía en cada proceso de hidrólisis.
5.- Describa el proceso de oxido-reducción.
Un gran número de reacciones químicas transcurre con la pérdida formal de electrones de un átomo y la ganancia de ellos por otro. La ganancia de electrones recibe el nombre de reducción y la pérdida de electrones oxidación. El proceso global se denomina oxido-reducción o reacción redox. La especie que suministra electrones es el agente reductor (especie que se oxida) y la que los gana es el agente oxidante (especie que se reduce). Estos hechos muestran que las reacciones redox se asemejan a las ácido-base según la definición de Brönsted, pero en lugar de transferirse protones desde un ácido a una base, en el caso de la oxido-reducción se transfieren electrones desde el agente reductor al oxidante.
6.- Mencione ¿Qué son los radicales libres?
Los radicales libres pueden formarse cuando se forman los enlaces covalentes, de tal forma que cada porción se queda con una mitad de los electrones compartidos; también se genera cuando un átomo o molécula acepte un solo electrón transferido durante una reacción de oxido reducción.
Los radicales libres son en extremo reactivos y pueden alterar de manera química muchos tipos de moléculas, entre ellas proteínas, lípidos y ácidos nucleicos. La formación de radicales hidroxilo tal vez sea una de las razones principales de que la luz solar sea tan nociva para la piel.
7.-Mencione brevemente la teoría quimiosmótica
La teoría sugiere esencialmente que la mayor parte de la síntesis de ATP en la respiración celular, viene de un gradiente electroquímico existente entre la membrana interna y el espacio intermembrana de la mitocondria, mediante el uso de la energía de NADH y FADH2 que se han formado por la ruptura de moléculas ricas en energía, como la glucosa.
8.- Esquematice como se lleva acabo la quimiosmosis en una mitocondria

9.- ¿Como se genera la fuerza protón motriz?
la FPM es generada por una cadena transportadora de electrones, que actúa como una bomba de electrones tanto como una de protones, bombeando electrones en direcciones opuestas, creando una separación de carga. En la mitocondria, la liberación de energía libre desde la cadena transportadora de electrones, es utilizada para mover protones desde la matriz mitocondrial al espacio intermembrana de la mitocondria. Mover los protones a las partes externas crea una concentraciones levemente alta de partículas cargadas positivamente, dando por resultado un lado más positivo y un lado más negativo (El gradiente eléctrico generado es de aproximadamente -200 mV (matriz negativa) )
Ésta diferencia de carga da como resultado un gradiente electroquímico. Éste gradiente se compone tanto del gradiente de pH y del gradiente eléctrico. El gradiente de pH corresponde a la diferencia en la concentración del ión H+. Juntos, el gradiente electroquímico de protones corresponde tanto a la concentración como a la diferencia de carga que se pueda generar, se denomina Fuerza Protón-Motriz (FPM).
10.- ¿De que manera es absorbida la luz a través de los fotosistemas?
La luz es absorbida por los pigmentos de la antena de fotosistemas II y I. La energía absorbida se transfiere a las clorofilas del Centro de Reacción, P 680 en el fotosistema II, P 700 en el fotosistema I. Absorción de 1 fotón de luz por el fotosistema II elimina 1 electrón de P 680. Con su carga positiva resultante, P 680 es lo suficientemente electronegativo que se puede quitar 1 electrón de una molécula de agua.Cuando estos pasos se han producido 4 veces, lo que requiere 2 moléculas de agua, 1 molécula de oxígeno y 4 de protones (H +) son liberados. Los electrones son transferidos (a través de plastoquinona - PQ en la figura) a la citocromo b 6 / f complejas en las que proporcionan la energía para quimiosmosis. La activación de P 700 en el fotosistema que le permite recoger los electrones del citocromo b 6 / f complejos (por medio de plastocianina - PC en la figura) y elevarlas a un potencial redox suficientemente alto para que, después de pasar por la ferredoxina, que pueden reducir NADP + a NADPH.
11.- ¿Como se lleva acabo la cadena de transporte de electrones fotosintéticos.?
En la fosforilación oxidativa, los electrones son transferidos desde un donador de electrones de alta energía a un aceptor a través de una cadena de transporte de electrones. En la fotofosforilación, la energía de la luz solar es usada para crear un donador de electrones altamente energético y un aceptor de esos electrones. Los electrones son transferidos desde el donador hasta el aceptor por una cadena de transporte totalmente diferente a la observada en las mitocondrias. La cadena de transporte de electrones fotosintética tiene varias similitudes con la cadena oxidativa. Tienen transportadores móviles, transportadores liposolubles y móviles, transportadores hidrosolubles y bombas de protones, que se encargan de generar el gradiente electroquímico.
12.- De que manera podemos apreciar los fenómenos fotobiólogicos
El sol provee, en forma de radiación electromagnética, la energía necesaria para el funcionamiento de todos los ecosistemas terrestres y acuáticos, basados en la producción de materia orgánica a través del proceso de fotosíntesis. La radiación solar, por lo tanto, es determinante para regular los ciclos naturales (estaciones, día-noche) así como otros factores (gradientes en altura y latitudinales de la vegetación, distribución vertical en la columna de agua, etc.). Sin embargo, esta misma radiación que hace posible la vida sobre la Tierra puede ser en algunos casos perjudiciales especialmente aquella parte de la radiación llamada radiación ultravioleta (RUV, 280-400 nm). Los organismos que fotosintetizan son los primeros en ser afectados, ya que ellos reciben la radiación solar y representan el primer punto de la red trófica.
13.- ¿Cuales son las características del Olfato?
El sentido del olfato nos permite recibir del exterior los estímulos olorosos. Este sentido, en general, está mucho más desarrollado en algunos animales que en el hombre.
En el hombre el sentido del olfato se localiza en una mucosa llamada mucosa olfativa, que se encuentra en el fondo de las fosas nasales. La mucosa olfativa está conectada a un nervio que transmite el estímulo hasta el cerebro llamado nervio olfatorio. En la parte superior y posterior de las fosas nasales está la mucosa pituitaria u olfativa, donde se perciben ciertos olores.
Igual que sucede en el caso de los sabores, existe un número limitado de olores que la mucosa olfativa puede percibir. En este caso, el número de olores primarios se ha calculado en siete, a cada uno de los cuales corresponde un perceptor olfativo particular.
14.- ¿Qué pasa cuando percibimos un olor?
automáticamente se estimulan los cilios, que empiezan un proceso de señales nerviosas, estás señales llegan al nervio olfativo y éste transmite las señales al bulbo olfativo. El cerebro es el encargado de identificar qué clase de olor estamos percibiendo, a través del proceso anterior.
Las neuronas de la región olfatoria son neuronas sensitivas primarias y forman parte de las neuronas cerebrales.
15.- ¿Qué trastornos nos producen perdida del olfato?
A menudo se pierde el sentido del olfato con trastornos que impiden que el aire llegue hasta el área de la nariz donde se localizan los receptores olfativos (la placa cribriforme ubicada en la parte superior de la nariz). Dichos trastornos pueden abarcar pólipos nasales, deformidades del tabique nasal y tumores nasales.
Otros trastornos que pueden producir una pérdida del sentido del olfato abarcan:
• Demencia de tipo Alzheimer
• Trastornos del sistema endocrino
• Traumatismo craneal
• Trastornos neurológicos
• Trastornos nutricionales
• Tumores de la cabeza o el cerebro.
16.- ¿Cuál es la función del sentido del gusto?
El gusto es función de las papilas gustativas en la boca; su importancia depende de que permita seleccionar los alimentos y bebidas según los deseos de la persona y también según las necesidades nutritivas. El sentido del gusto depende de la estimulación de los llamados “botones gustativos”, las cuales se sitúan preferentemente en la lengua, aunque algunas se encuentran en el paladar; su sensibilidad es variable.
17.-¿ cuales son los 4 sabores elementales que podemos percibir?
La sensación denominada dulce.
•La sensación denominada ácida
•La sensación denominada salada
•La sensación denominada amarga
18.- Mencione que producen los trastornos del sentido del gusto.
Las alteraciones en el sentido del gusto son problemas que por lo general han recibido poca atención de la comunidad médica, por una parte el diagnóstico es complicado, y por otra no existe un tratamiento estandarizado. Se hace una revisión de la literatura y se enfatiza la necesidad del estomatólogo para profundizar sobre estos trastornos, para formar parte del equipo multidisciplinario que se requiere en este tipo de pacientes.
19.- de que manera llegan los rayos luminosos a nuestros ojos
Los rayos luminosos son captados por las células receptoras de la retina y conducidos hacia el nervio óptico, que nace en la parte posterior de la capa sensorial. Después de pasar por el quiasma óptico, donde una parte de sus fibras se entrecruzan, forman las cintillas ópticas, que llegan al lóbulo occipital del cerebro.
20.- como se da el proceso de la visión
El principal estímulo de la vista es la luz y el campo receptor es la retina.
La luz pasa a través de:
La córnea y el humor acuoso (primera refracción)
La pupila, el cristalino (segunda refracción)
Humor vítreo, retina (tercera refracción)
Descripción:
La córnea es, una estructura transparente y constituye el elemento refractivo principal del ojo.
El humor acuoso es, un líquido claro que ocupa el espacio entre la córnea y el cristalino.
La pupila es, la abertura dilatable y contráctil en el centro del iris por la que pasan los rayos luminosos.
El cristalino es, un cuerpo lenticular, transparente, situado entre el humor acuoso y el cuerpo vítreo.
Humor vítreo es, la sustancia que llena la cámara posterior del ojo, detrás del cristalino.
La retina es, la membrana más interna de las tres que forman el globo ocular.
El globo ocular está protegido dentro de la órbita y solamente está expuesta la sexta parte. La parte expuesta está protegida por los párpados. Las glándulas lacrimales producen lágrimas para limpiar, facilitar el deslizamiento de los párpados y humectar a las células de la conjuntiva.
El ojo debe realizar dos ajustes uno para la intensidad de la luz y otro para la distancia.
El iris (es la parte pigmentada o color del ojo) puede abrirse o encogerse cuando hay poca luz o cerrarse cuando hay demasiada luz, que consta de fibras musculares radiales y circulares.
Es el análisis cuantitativo de la forma en que los organismos adquieren y utilizan la energía, la bioenergética puede considerase una parte especial de la ciencia de las transformaciones energéticas denominada termodinámica.
2.- ¿Qué es un sistema termodinámico?
Un sistema termodinámico, es la parte del universo que elegimos para el estudio, tal como un recipiente de reacción, un organismo, una única célula bacteriana, el resto del universo se considera como el entorno, este debe tener unos límites definidos.
3.- ¿Por qué el ATP es muy importante en los sistemas vivos?
Las células requieren un continuo suministro de energía. Esta es necesaria para la síntesis de moléculas complejas, la ejecución de trabajo mecánico y el transporte de sustancias a través de sus membranas. La energía es transferida desde las reacciones químicas que la acumulan a las que las consumen, mediante una molécula especial, el ATP. En las células, la energía que recibe o cede el ATP es la contenida en el enlace entre su último fosfato y el resto de la molécula. El enlace se forma durante la síntesis de ATP: incorporan así energía, la que se cede cuando el enlace se escinde.
4.- ¿Mencione la relación que hay entre el ATP y la energía libre?
En la hidrólisis del fosfoenolpiruvato, el bifosfoglicerato , el adenosina trifosfato y el pirofosfato, los dos productos de la hidrólisis llevan una carga negativa.
La repulsión entre estos productos iónicos favorece intensamente la reacción de hidrólisis
La hidrólisis se ve favorecida a veces no sólo por la estabilización de la resonancia del ortofosfato sino también por la capacidad del otro producto para adoptar más formas moleculares, lo cual contribuye a la hidrólisis del del fosfoenolpiruvato ya que el piruvato puede tautomerizarse, con la liberación de energía en cada proceso de hidrólisis.
5.- Describa el proceso de oxido-reducción.
Un gran número de reacciones químicas transcurre con la pérdida formal de electrones de un átomo y la ganancia de ellos por otro. La ganancia de electrones recibe el nombre de reducción y la pérdida de electrones oxidación. El proceso global se denomina oxido-reducción o reacción redox. La especie que suministra electrones es el agente reductor (especie que se oxida) y la que los gana es el agente oxidante (especie que se reduce). Estos hechos muestran que las reacciones redox se asemejan a las ácido-base según la definición de Brönsted, pero en lugar de transferirse protones desde un ácido a una base, en el caso de la oxido-reducción se transfieren electrones desde el agente reductor al oxidante.
6.- Mencione ¿Qué son los radicales libres?
Los radicales libres pueden formarse cuando se forman los enlaces covalentes, de tal forma que cada porción se queda con una mitad de los electrones compartidos; también se genera cuando un átomo o molécula acepte un solo electrón transferido durante una reacción de oxido reducción.
Los radicales libres son en extremo reactivos y pueden alterar de manera química muchos tipos de moléculas, entre ellas proteínas, lípidos y ácidos nucleicos. La formación de radicales hidroxilo tal vez sea una de las razones principales de que la luz solar sea tan nociva para la piel.
7.-Mencione brevemente la teoría quimiosmótica
La teoría sugiere esencialmente que la mayor parte de la síntesis de ATP en la respiración celular, viene de un gradiente electroquímico existente entre la membrana interna y el espacio intermembrana de la mitocondria, mediante el uso de la energía de NADH y FADH2 que se han formado por la ruptura de moléculas ricas en energía, como la glucosa.
8.- Esquematice como se lleva acabo la quimiosmosis en una mitocondria

9.- ¿Como se genera la fuerza protón motriz?
la FPM es generada por una cadena transportadora de electrones, que actúa como una bomba de electrones tanto como una de protones, bombeando electrones en direcciones opuestas, creando una separación de carga. En la mitocondria, la liberación de energía libre desde la cadena transportadora de electrones, es utilizada para mover protones desde la matriz mitocondrial al espacio intermembrana de la mitocondria. Mover los protones a las partes externas crea una concentraciones levemente alta de partículas cargadas positivamente, dando por resultado un lado más positivo y un lado más negativo (El gradiente eléctrico generado es de aproximadamente -200 mV (matriz negativa) )
Ésta diferencia de carga da como resultado un gradiente electroquímico. Éste gradiente se compone tanto del gradiente de pH y del gradiente eléctrico. El gradiente de pH corresponde a la diferencia en la concentración del ión H+. Juntos, el gradiente electroquímico de protones corresponde tanto a la concentración como a la diferencia de carga que se pueda generar, se denomina Fuerza Protón-Motriz (FPM).
10.- ¿De que manera es absorbida la luz a través de los fotosistemas?
La luz es absorbida por los pigmentos de la antena de fotosistemas II y I. La energía absorbida se transfiere a las clorofilas del Centro de Reacción, P 680 en el fotosistema II, P 700 en el fotosistema I. Absorción de 1 fotón de luz por el fotosistema II elimina 1 electrón de P 680. Con su carga positiva resultante, P 680 es lo suficientemente electronegativo que se puede quitar 1 electrón de una molécula de agua.Cuando estos pasos se han producido 4 veces, lo que requiere 2 moléculas de agua, 1 molécula de oxígeno y 4 de protones (H +) son liberados. Los electrones son transferidos (a través de plastoquinona - PQ en la figura) a la citocromo b 6 / f complejas en las que proporcionan la energía para quimiosmosis. La activación de P 700 en el fotosistema que le permite recoger los electrones del citocromo b 6 / f complejos (por medio de plastocianina - PC en la figura) y elevarlas a un potencial redox suficientemente alto para que, después de pasar por la ferredoxina, que pueden reducir NADP + a NADPH.
11.- ¿Como se lleva acabo la cadena de transporte de electrones fotosintéticos.?
En la fosforilación oxidativa, los electrones son transferidos desde un donador de electrones de alta energía a un aceptor a través de una cadena de transporte de electrones. En la fotofosforilación, la energía de la luz solar es usada para crear un donador de electrones altamente energético y un aceptor de esos electrones. Los electrones son transferidos desde el donador hasta el aceptor por una cadena de transporte totalmente diferente a la observada en las mitocondrias. La cadena de transporte de electrones fotosintética tiene varias similitudes con la cadena oxidativa. Tienen transportadores móviles, transportadores liposolubles y móviles, transportadores hidrosolubles y bombas de protones, que se encargan de generar el gradiente electroquímico.
12.- De que manera podemos apreciar los fenómenos fotobiólogicos
El sol provee, en forma de radiación electromagnética, la energía necesaria para el funcionamiento de todos los ecosistemas terrestres y acuáticos, basados en la producción de materia orgánica a través del proceso de fotosíntesis. La radiación solar, por lo tanto, es determinante para regular los ciclos naturales (estaciones, día-noche) así como otros factores (gradientes en altura y latitudinales de la vegetación, distribución vertical en la columna de agua, etc.). Sin embargo, esta misma radiación que hace posible la vida sobre la Tierra puede ser en algunos casos perjudiciales especialmente aquella parte de la radiación llamada radiación ultravioleta (RUV, 280-400 nm). Los organismos que fotosintetizan son los primeros en ser afectados, ya que ellos reciben la radiación solar y representan el primer punto de la red trófica.
13.- ¿Cuales son las características del Olfato?
El sentido del olfato nos permite recibir del exterior los estímulos olorosos. Este sentido, en general, está mucho más desarrollado en algunos animales que en el hombre.
En el hombre el sentido del olfato se localiza en una mucosa llamada mucosa olfativa, que se encuentra en el fondo de las fosas nasales. La mucosa olfativa está conectada a un nervio que transmite el estímulo hasta el cerebro llamado nervio olfatorio. En la parte superior y posterior de las fosas nasales está la mucosa pituitaria u olfativa, donde se perciben ciertos olores.
Igual que sucede en el caso de los sabores, existe un número limitado de olores que la mucosa olfativa puede percibir. En este caso, el número de olores primarios se ha calculado en siete, a cada uno de los cuales corresponde un perceptor olfativo particular.
14.- ¿Qué pasa cuando percibimos un olor?
automáticamente se estimulan los cilios, que empiezan un proceso de señales nerviosas, estás señales llegan al nervio olfativo y éste transmite las señales al bulbo olfativo. El cerebro es el encargado de identificar qué clase de olor estamos percibiendo, a través del proceso anterior.
Las neuronas de la región olfatoria son neuronas sensitivas primarias y forman parte de las neuronas cerebrales.
15.- ¿Qué trastornos nos producen perdida del olfato?
A menudo se pierde el sentido del olfato con trastornos que impiden que el aire llegue hasta el área de la nariz donde se localizan los receptores olfativos (la placa cribriforme ubicada en la parte superior de la nariz). Dichos trastornos pueden abarcar pólipos nasales, deformidades del tabique nasal y tumores nasales.
Otros trastornos que pueden producir una pérdida del sentido del olfato abarcan:
• Demencia de tipo Alzheimer
• Trastornos del sistema endocrino
• Traumatismo craneal
• Trastornos neurológicos
• Trastornos nutricionales
• Tumores de la cabeza o el cerebro.
16.- ¿Cuál es la función del sentido del gusto?
El gusto es función de las papilas gustativas en la boca; su importancia depende de que permita seleccionar los alimentos y bebidas según los deseos de la persona y también según las necesidades nutritivas. El sentido del gusto depende de la estimulación de los llamados “botones gustativos”, las cuales se sitúan preferentemente en la lengua, aunque algunas se encuentran en el paladar; su sensibilidad es variable.
17.-¿ cuales son los 4 sabores elementales que podemos percibir?
La sensación denominada dulce.
•La sensación denominada ácida
•La sensación denominada salada
•La sensación denominada amarga
18.- Mencione que producen los trastornos del sentido del gusto.
Las alteraciones en el sentido del gusto son problemas que por lo general han recibido poca atención de la comunidad médica, por una parte el diagnóstico es complicado, y por otra no existe un tratamiento estandarizado. Se hace una revisión de la literatura y se enfatiza la necesidad del estomatólogo para profundizar sobre estos trastornos, para formar parte del equipo multidisciplinario que se requiere en este tipo de pacientes.
19.- de que manera llegan los rayos luminosos a nuestros ojos
Los rayos luminosos son captados por las células receptoras de la retina y conducidos hacia el nervio óptico, que nace en la parte posterior de la capa sensorial. Después de pasar por el quiasma óptico, donde una parte de sus fibras se entrecruzan, forman las cintillas ópticas, que llegan al lóbulo occipital del cerebro.
20.- como se da el proceso de la visión
El principal estímulo de la vista es la luz y el campo receptor es la retina.
La luz pasa a través de:
La córnea y el humor acuoso (primera refracción)
La pupila, el cristalino (segunda refracción)
Humor vítreo, retina (tercera refracción)
Descripción:
La córnea es, una estructura transparente y constituye el elemento refractivo principal del ojo.
El humor acuoso es, un líquido claro que ocupa el espacio entre la córnea y el cristalino.
La pupila es, la abertura dilatable y contráctil en el centro del iris por la que pasan los rayos luminosos.
El cristalino es, un cuerpo lenticular, transparente, situado entre el humor acuoso y el cuerpo vítreo.
Humor vítreo es, la sustancia que llena la cámara posterior del ojo, detrás del cristalino.
La retina es, la membrana más interna de las tres que forman el globo ocular.
El globo ocular está protegido dentro de la órbita y solamente está expuesta la sexta parte. La parte expuesta está protegida por los párpados. Las glándulas lacrimales producen lágrimas para limpiar, facilitar el deslizamiento de los párpados y humectar a las células de la conjuntiva.
El ojo debe realizar dos ajustes uno para la intensidad de la luz y otro para la distancia.
El iris (es la parte pigmentada o color del ojo) puede abrirse o encogerse cuando hay poca luz o cerrarse cuando hay demasiada luz, que consta de fibras musculares radiales y circulares.
martes, 30 de marzo de 2010
OLFATO

En el hombre el sentido del olfato se localiza en una mucosa llamada mucosa olfativa, que se encuentra en el fondo de las fosas nasales. La mucosa olfativa está conectada a un nervio que transmite el estímulo hasta el cerebro llamado nervio olfatorio. En la parte superior y posterior de las fosas nasales está la mucosa pituitaria u olfativa, donde se perciben ciertos olores.
Igual que sucede en el caso de los sabores, existe un número limitado de olores que la mucosa olfativa puede percibir. En este caso, el número de olores primarios se ha calculado en siete, a cada uno de los cuales corresponde un perceptor olfativo particular.

El sentido del olfato está muy relacionado con el del gusto. En efecto, tú mismo habrás podido comprobar que cuando estás acatarrado la comida parece tener menos sabor. Esta diferencia se debe a que en ese momento el sentido del olfato permanece muy amortiguado, lo que también afecta a la percepción de los sabores.
Estos dos últimos sentidos, el del gusto y el olfativo, están estrechamente relacionado, por eso cuando una persona está resfriada no distingue correctamente los sabores.
Detrás de los agujeros de la nariz se encuentra el epitelio olfativo, el cual está formado por un grupo de células nerviosas cubiertas de unos finísimos pelitos, llamados cilios, y a su vez los cilios están cubiertos por unos receptores sensibles a las moléculas del olor.
Existen por lo menos 20 diferentes tipos de receptores y cada uno tiene la capacidad de sentir una determinada clase de moléculas de olor.
Cuando percibimos un olor, automáticamente se estimulan los cilios, que empiezan un proceso de señales nerviosas, estás señales llegan al nervio olfativo y éste transmite las señales al bulbo olfativo. El cerebro es el encargado de identificar qué clase de olor estamos percibiendo, a través del proceso anterior.
Las neuronas de la región olfatoria son neuronas sensitivas primarias y forman parte de las neuronas cerebrales.
Como hemos visto el proceso es complicado y sorprendente, ya que toda la acción se realiza en fracciones de segundo.
El sentido del olfato es capaz de desencadenar en nuestro organismo reacciones como: abrir el apetito, eliminar el apetito, recuerdos de ciertos momentos o personas especiales, reacciones físicas y muchas más.

Es común que se presente la pérdida temporal del sentido del olfato con los resfriados comunes y las alergias nasales, como la fiebre del heno (rinitis alérgica), al igual que después de una enfermedad viral.
Algo de pérdida del olfato se presenta al envejecer. En la mayoría de los casos, no hay una causa obvia o inmediata y no hay un tratamiento.
A menudo se pierde el sentido del olfato con trastornos que impiden que el aire llegue hasta el área de la nariz donde se localizan los receptores olfativos (la placa cribriforme ubicada en la parte superior de la nariz). Dichos trastornos pueden abarcar pólipos nasales, deformidades del tabique nasal y tumores nasales.
Otros trastornos que pueden producir una pérdida del sentido del olfato abarcan:
• Demencia de tipo Alzheimer
• Trastornos del sistema endocrino
• Traumatismo craneal
• Trastornos neurológicos
• Trastornos nutricionales
• Tumores de la cabeza o el cerebro
GUSTO


Los nervios conectados con las papilas gustativas transmiten impulsos al centro nervioso situado en el bulbo raquídeo (continuación de la médula allí donde empieza la columna vertebral); de aquí, los impulsos se transmiten a las caras superior e interna del lóbulo temporal, en íntima relación con el área del cerebro relacionada con el olfato.
A partir de los estudios psicológicos, se piensa en general que existen cuando menos cuatro sensaciones sápidas primarias: ácido, salado, dulce y amargo; pero sabemos que una persona puede percibir cientos o miles de sabores diferentes. Se supone que se trata de combinaciones de las cuatro sensaciones primarias, de la misma manera que todos los colores del espectro son combinaciones de tres sensaciones coloreadas primarias. Sin embargo, podría existir otra clase o subclase de sensaciones primarias, menos evidentes.
Sensaciones sápidas primarias
Los fisiólogos han identificado los cuatro sabores elementales y los han codificado en los siguientes términos:
•La sensación denominada dulce.
•La sensación denominada ácida
•La sensación denominada salada
•La sensación denominada amarga
Sobre la lengua, las zonas de percepción de cada uno de los sabores son netamente diferenciados. El borde lateral derecho e izquierdo de la lengua reconoce las sensaciones saladas (1); las zonas situadas en el extremo de la lengua reconoce el sabor dulce(2). El amargo es percibido por un sector al fondo de la lengua que tiene la forma de acento circunflejo (3). La acidez es percibida por los botones o papilas situadas a los costados de la lengua por debajo de las papilas que perciben los sabores salados (4).

Sabor ácido.
Está causado por ácidos, y la intensidad de la sensación gustativa es aproximadamente proporcional a la concentración de iones hidrógeno. En otras palabras, cuanto más fuerte es el ácido, más intensa la sensación.
Se puede reconocer este sabor, agregando una gotas de ácido orgánico natural, como el cítrico a un vaso de agua.
Esta sensación afecta las zonas laterales de la lengua, por debajo de la zona donde se perciben los sabores salados. Este sabor irrita ligeramente las mucosas y se produce secreción de gran cantidad de saliva bien fluida.
Este sabor es fácil de reconocer, porque se asocia a los frutos verdes o al vinagre.
El vino es rico en diversos ácidos, que en su conjunto otorgan la característica ácida en una amplia gama.
Sabor salado.
El gusto salado depende de sales ionizadas. La calidad del gusto varía algo de una sal a otra, porque las sales también estimulan otros botones gustativos en grado variable.
Si a un poco de agua le agregamos un poco de sal de cocina, percibimos una sensación particular, sobre todo en los bordes laterales de la lengua, que es acompañado por una secreción fugaz de saliva.
Este sabor es casi inexistente en los vinos, pero no se debe ignorar
Sabor dulce.
No depende de ninguna clase aislada de productos químicos. Una lista de algunos productos químicos que causan este sabor es la siguiente: azúcares, glicoles, alcoholes, aldehídos, cetonas, amidas, ésteres, aminoácidos, etc. Obsérvese específicamente que casi todas las sustancias que causan sabor dulce son productos químicos orgánicos.
Si degustamos un vaso de agua al cual se le ha agregado azúcar alimentaria (sacarosa), se crea una impresión característica en la punta de la lengua que es la zona fundamental de reconocimiento de este sabor, también sobre los labios, la mucosa de la boca a nivel de las encías inferiores. Ellas hacen secretar una saliva espesa y viscosa. La mayor parte de los vinos son secos y no contienen azúcar (excepto algunos blancos y licorosos). Sin embargo, a veces se perciben de esta manera, sustancias cono las ya citadas, correspondientes al alcohol, glicerol o trazas de fructuosa y de pentosa.
Sabor amargo.
El sabor amargo, como el dulce no depende de un solo tipo de agente químico. Aquí también, las sustancias que dan sabor amargo son casi todas de tipo orgánico.
El sabor amargo puede percibirse particularmente en los vinos tintos aún sanos, por su riqueza polifenólica, sobre todo en taninos. Es conocido que los taninos tienen la particularidad de combinarse con las proteínas.
En los vinos tintos jóvenes, ricos en sustancias tánicas, estos cuerpos se combina con las proteínas de la saliva, secando la boca. Produciendo al mismo tiempo una sensación rasposa sobre dientes y encías. A veces en el fondo de la lengua dejan una sensación de astringencia.
A medida que el vino tinto madura y envejece, los taninos se van acomplejando más y más, para terminar suavizándose.
El ligero amargor de los taninos, que se pierde con el tiempo, no debe confundirse con el amargor de un vino enfermo por ataque bacteriano o su contenido en glicerol.
Así es como las sensaciones de astringencia y amargor no se revelan en los vinos blancos y rosados. Si ello ocurriera se debe a anomalías de carácter físico- químico y biológico, extrañas a la calidad elemental que debe caracterizar a los mismos.
En laboratorio se puede crear la sensación amarga con algunos miligramos de sal de quinina en un litro de agua.
Trastornos en el sentido del gusto.
Las alteraciones en el sentido del gusto son problemas que por lo general han recibido poca atención de la comunidad médica, por una parte el diagnóstico es complicado, y por otra no existe un tratamiento estandarizado. Se hace una revisión de la literatura y se enfatiza la necesidad del estomatólogo para profundizar sobre estos trastornos, para formar parte del equipo multidisciplinario que se requiere en este tipo de pacientes
lunes, 29 de marzo de 2010
VISION

El principal estímulo de la vista es la luz y el campo receptor es la retina.
La luz pasa a través de:
La córnea y el humor acuoso (primera refracción)
La pupila, el cristalino (segunda refracción)
Humor vítreo, retina (tercera refracción)
Descripción:
La córnea es, una estructura transparente y constituye el elemento refractivo principal del ojo.
El humor acuoso es, un líquido claro que ocupa el espacio entre la córnea y el cristalino.
La pupila es, la abertura dilatable y contráctil en el centro del iris por la que pasan los rayos luminosos.
El cristalino es, un cuerpo lenticular, transparente, situado entre el humor acuoso y el cuerpo vítreo.
Humor vítreo es, la sustancia que llena la cámara posterior del ojo, detrás del cristalino.
La retina es, la membrana más interna de las tres que forman el globo ocular.
El globo ocular está protegido dentro de la órbita y solamente está expuesta la sexta parte. La parte expuesta está protegida por los párpados. Las glándulas lacrimales producen lágrimas para limpiar, facilitar el deslizamiento de los párpados y humectar a las células de la conjuntiva.
El ojo debe realizar dos ajustes uno para la intensidad de la luz y otro para la distancia.
El iris (es la parte pigmentada o color del ojo) puede abrirse o encogerse cuando hay poca luz o cerrarse cuando hay demasiada luz, que consta de fibras musculares radiales y circulares.
La distancia la determina el cristalino, aumentando la curvatura de su cara anterior para la visión cercana y aplanándola para la visión lejana.
En concreto, la luz entra a través de la córnea, después, el rayo luminoso encuentra el iris, y a través de la pupila el rayo luminoso pasa al cristalino, después del cristalino, la luz atraviesa una masa gelatinosa clara, el humor vítreo, por último, el rayo de luz llega a la retina.
Las impresiones obtenidas por las células sensoriales de la retina son conducidas por el nervio óptico y posteriormente a la vía óptica, al centro visual del cerebro, donde la imagen toma forma y la percibimos.
Formación de las Imágenes

Para tener una idea clara del funcionamiento del ojo, se lo compara con una cámara fotográfica: el cristalino constituye el objetivo: el iris es un diafragma que regula la cantidad de luz agrandándose o achicándose; el humor vítreo es la cámara oscura; la coroides constituye las paredes de esta cámara, y la retina, la placa sensible.
Los rayos luminosos, después de atravesar el aparato dióptrico, excitan las sensoriales receptoras de la retina formando la imagen invertida del objeto que miramos. Esta inversión se debe a las distintas densidades de los medios que refractan la luz, de modo que los rayos luminosos superiores se proyectan en la parte inferior de la retina y los inferiores se dirigen a la parte superior.
En el centro de la visión, que se encuentra en la zona occipital del cerebro, donde la percepción se vuelve consciente, la imagen se endereza por un mecanismo aún desconocido, pero que se supone es psíquico.
Acomodación del ojo
El ojo sufre una doble acomodación. A la luz y a la distancia.
Acomodación a la luz. Esta función la cumple el iris, achicando la pupila cuando la intensidad luminosa es mayor, y agrandándola cuando la intensidad es menor. Esta acción está determinada por los músculos circulares y radiales.
Acomodación a la distancia. Esta función está determinada por el cristalino, aumentando la curvatura de su cara anterior, para la visión cercana y aplanándola para la visión lejana.
Visión Binocular
Consiste en la observación de un campo visual con los dos ojos al mismo tiempo. La visión de las dos imágenes formadas en ambas retinas se superponen y el cerebro interpreta una sola imagen más completa y precisa.
Persistencia Retiniana
Las imágenes persisten un breve tiempo en la retina.
El ojo humano no puede separar más de 10 imágenes por segundo. De esta imperfección se valen la televisión y la cinematografía, que proyectan más de 20 imágenes por segundo; si se pasaran 6 imágenes por segundo, apreciaríamos las diferencias entre las diversas posiciones. La sucesión rápida de imágenes nos da la impresión de movimiento.
Ilusiones Opticas
Reciben este nombre los errores que cometemos al emitir juicios visuales. Por ejemplo, cuando observamos dos círculos iguales y uno de ellos nos parece mayor.


Vía Optica
Es el camino que siguen las impresiones visuales desde la retina hasta el centro de la visión, ubicado en la zona occipital de los hemisferios cerebrales


Los rayos luminosos son captados por las células receptoras de la retina y conducidos hacia el nervio óptico, que nace en la parte posterior de la capa sensorial. Después de pasar por el quiasma óptico, donde una parte de sus fibras se entrecruzan, forman las cintillas ópticas, que llegan al lóbulo occipital del cerebro.
Anomalías del Ojo
Cuando las imágenes se forman en la retina (fig. A) se dice que el ojo es normal o emétrope (del friego em, en y métrom, medida).
En otros casos la imagen se forma por delante o por detrás de la retina; se dice entonces que el ojo es amétrope (del griego a, sin y métron, medida). Si la imagen se forma por delante de la retina(fig. B), el ojo se denomina miope (del griego myein, cerrar y ops, ojo); y si se forma por detrás, se dice que es hipermétrope (del griego hyper, más allá y métron, medida).
La miopía se produce cuando el diámetro anteroposterior del globo ocular es demasiado largo(fig. B), o cuando la cara anterior del cristalino es demasiado convexa(fig. C). Los que padecen de esta anomalía no ven bien a la distancia, y necesitan lentes bicóncavas. (1)
La hipermetropía se presenta cuando el diámetro anteroposterior del globo ocular es corto(fig. C) o cuando la cara anterior del cristalino es casi plana(fig. D). El hipermétrope no ve bien de cerca y su defecto se corrige con lentes biconvexas. (b)
Astigmatismo (del griego a, sin y stigma, punto). La córnea presenta una desigual covertura en los distintos planos, deformando las imágenes; por ejemplo, en un principio la cifra 42 se convierte en 24. Esta anomalía se corrige con lentes cilíndricas pulidas en forma despareja para compensar los desniveles de la córnea.
Presbicia, esta anomalía es común en los ancianos y se produce por la pérdida de elasticidad del cristalino, que no sufre acomodación.
Estrabismo, es un defecto de las contracciones de los músculos del ojo, a causa del cual la visión no es perfecta.
Daltonismo, consiste en la confusión de los colores rojo y verde. Recibe este nombre en honor del físico inglés Dalton, que padeció este trastorno visual y que lo describió en el año 1798.
El daltonismo es hereditario, y bastante frecuente. Las personas que padecen de este mal no pueden realizar trabajos en los que se usen señales coloreadas (maquinistas, conductores, etc).
Acromatopsia, es la ceguera completa para los colores. Se debe a que ninguna de las fibras nerviosas es excitable por éstos. Esta enfermedad es poco frecuente.
Catarata, esta enfermedad característica de la vejez, consiste en el enturbamiento del cristalino como consecuencia de una coagulación.
La luz, que llega a nuestros ojos y nos permite ver, es un pequeño conjunto de radiaciones electromagnéticas de longitudes de onda comprendidas entre los 380 nm y los 770 nm.
El espectro electromagnético
La luz forma parte del espectro electromagnético que comprende tipos de ondas tan dispares como los rayos cósmicos, los rayos gamma, los ultravioletas, los infrarrojos y las ondas de radio o televisión entre otros. Cada uno de estos tipos de onda comprende un intervalo definido por una magnitud característica que puede ser la longitud de onda ( ) o la frecuencia (f). Recordemos que la relación entre ambas es:
donde c es la velocidad de la luz en el vacío (c = 3•108 m/s).

TRANSPORTE ATRAVEZ DE MEMBRANAS
Movimiento del agua y los solutos
La membrana celular actúa como barrera semipermeable impidiendo la entrada de la mayor parte de las moléculas, dejando pasar selectivamente a otras.
Para entender los sucesos que acontecen es necesario refrescar los conceptos de
El potencial de agua es la tendencia del agua a moverse de un área de mayor concentración a una de menor concentración. Las moléculas de agua se mueven de acuerdo a la diferencia de energía potencial entre el punto donde se encuentran y el lugar hacia donde se dirigen. La presión y la gravedad son dos de los orígenes de este movimiento.
Recuerde por ejemplo el ciclo hidrológico en el cual el agua fluye de las partes altas a las bajas, al igual que el agua de lluvia cae de las nubes, y para volver a formar parte de ellas es necesario que el sol la evapore. La energía es necesaria tanto para mantener este ciclo, como para llevar el agua a una zona alta.
Difusión simple

Células y Difusión

Para que tenga lugar el fenómeno de la difusión, la distribución espacial de moléculas no debe ser homogénea, debe existir una diferencia, o gradiente de concentración entre dos puntos del medio.
La difusión constituye una de las principales formas de movimiento de sustancias entre las células y una de las formas en que las pequeñas moléculas cruzan la membrana celular.
El intercambio de gases en branquias y pulmones es consecuencia de fenómenos de difusión. El anhídrido carbónico se regenera constantemente dado que es producido en las células como consecuencia de fenómenos metabólicos, y como la fuente está en el interior de la célula, el flujo neto del CO2 es hacia el exterior de la célula. Los procesos metabólicos, requieren usualmente oxígeno, cuya concentración es mayor en el exterior de la célula, por lo tanto su flujo neto es hacia el interior.
El primer paso de la difusión pasiva en el entorno celular, es el movimiento de una molécula desde la solución acuosa a la porción hidrofóbica de la bicapa fosfolipídica.

La medida de la "fobia" al agua de una molécula, esta dada por su coeficiente de partición K.

K es una medida de la tendencia de una molécula a "huir" del agua, a mayor K, mayor es la posibilidad de disolución de una molécula en la bicapa lipídica.
La velocidad de difusión (dn/dt expresada en mol/s) de moléculas pequeñas a través de una membrana esta dada por una variante de la Ley de Fick y de acuerdo a ella, la misma es proporcional a la diferencia de concentraciones a ambos lados de la membrana (C1aq-C2aq). Además, para una membrana de superficie A y de espesor x, es proporcional a A e inversamente proporcional a x. Por otra parte resulta proporcional a P (coeficiente de permeabilidad), todo lo cual resulta en:
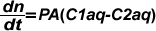



Ósmosis
Ejemplos de ese tipo de membrana son la membrana celular, como así también productos como los tubos de diálisis y las envolturas de acetato de celulosa de algunas salchichas.
La presencia de solutos decrece el potencial de agua de una sustancia, por lo tanto existe más agua por unidad de volumen en un vaso de agua corriente que en el volumen equivalente de agua de mar.
En una célula, que posee organelas y moléculas grandes, la dirección del flujo del agua es, generalmente, hacia el interior de la célula.
La presión osmótica se define como la presión hidrostática necesaria para detener el flujo neto de agua a través de una membrana semipermeable que separa soluciones de composición diferente. La presión osmótica (p) está dada por:

La presión osmótica es una propiedad de tipo coligativa, es decir, depende del número de partículas. Así por ejemplo una solución de NaCl 0,5 M, si estuviera totalmente disociada en Na+ y Cl-, sería equivalente a una solución de glucosa 1M.
Las soluciones hipertónicas son aquellas, que con referencias al interior de la célula, contienen mayor cantidad de solutos (y por lo tanto menor potencial de agua).
Las hipotónicas son aquellas, que en cambio contienen menor cantidad de solutos (o, en otras palabras, mayor potencial de agua).
Las soluciones isotónicas tienen concentraciones equivalentes de solutos y, en este caso, al existir igual cantidad de movimiento de agua hacia y desde el exterior, el flujo neto es nulo.

Una de las principales funciones del cuerpo de los animales es el mantenimiento de la isotonicidad del plasma sanguíneo, es decir un medio interno isotónico. Esto elimina los problemas asociados con la pérdida o ganancia de agua desde y hacia las células. Estamos hablando por supuesto de una de las claves de la homeostasis.
A diferencia de la células animales, las células de bacterias y plantas están rodeadas por una pared celular rígida, en este caso
Cuando se encuentran en un medio hipotónico, el agua que penetra por flujo osmótico genera una presión de turgencia que empuja al citosol y la membrana plasmática contra la pared celular.
En cambio en soluciones hipertónicas las células se retraen, separándose la membrana de la pared celular como consecuencia de la pérdida de agua por flujo osmótico (fenómeno conocido como plasmólisis).
Organismos unicelulares como Paramecium, y otros organismos de vida libre en agua dulce, tienen el problema de que son usualmente hipertónicos con relación a su medio ambiente. Por lo tanto el agua tiende a fluir a través de la membrana hinchando a la célula y eventualmente rompiéndola, hecho molesto para cualquier célula. Una vacuola contráctil es la respuesta del Paramecium a este problema, si bien el bombear agua hacia exterior de la célula requiere energía ya que trabaja contra un gradiente de concentración.
Proteínas de membrana que intervienen en el transporte
El transporte de moléculas es realizado por parte de las proteínas integradas en la membrana celular. Por lo general es altamente selectivo en lo que se refiere a los productos químicos que permiten pasar.
Las tres clases principales de proteínas de membrana (todas ellas de transmembrana) que intervienen en el pasaje de moléculas a través de la misma son:
proteínas de canal que conforman un "túnel" que permite el paso de agua y electrolitos a favor de un gradiente de concentración o potencial eléctrico (forman un canal que atraviesa la bicapa en todo su espesor). La partícula que pasa se selecciona de acuerdo a su tamaño y carga. Suelen estar cerrados y abrirse frente a estímulos específicos. El pasaje se realiza de acuerdo al gradiente de concentración de las moléculas.
Las células que presentan gran permeabilidad al agua poseen un canal que facilita la entrada de la misma. La proteína responsable: la acuoporina, fue identificada por Peter Agre en eritrocitos, a mediados de los ´80.

¿Cómo trabaja el canal de agua ? Una pregunta de que llevó al NOBEL
En 2000, junto con otros equipos de investigación, Agre informó las primeras imágenes de la estructura tridimensional de la aquaporina. Con estos datos, era posible trazar en detalle cómo funciona el canal de agua. ¿Porque sólo admite las moléculas de agua y no otras moléculas o iones? por ejemplo, no permite que pasen los protones. Esto es crucial porque la diferencia en la concentración de protones entre el interior y el exterior de la célula es la base del sistema de almacenamiento de energía de la célula.
La selectividad es una propiedad central de la aquaporina. Debido a la carga positiva del centro del canal, los iones cargados positivamente se rechazan. Esto impide el pasaje de protones a través del mismo. Las moléculas que se introducen a través del estrecho canal se reacomodan, orientándose en el campo eléctrico local formado por los átomos de la pared. Los protones (o más bien los iones oxonium, H3O+) se detienen en el camino y son rechazados debido a sus cargas positivas.



Proteína transportadora
sufren un cambio de conformación (B) en manera tal que permite a las moléculas fijadas, atravesar la membrana plasmática. Se conocen tres tipos de transportadores:

Tipos de transportadores
"uniport" llevan una soluto por vez
"symport" transportan el soluto y co-transportan otro diferente al mismo tiempo y en la misma dirección.
"antiport" transportan soluto hacia el interior (o exterior) y co-transportan soluto en la dirección opuesta. Uno entra y el otro sale o vice-versa.
El transporte activo, en cambio, requiere por parte de la célula un gasto de energía que usualmente se da en la forma de consumo de ATP. Ejemplos del mismo son el transporte de moléculas de gran tamaño (no solubles en lípidos) y la bomba sodio-potasio.
Difusión facilitada
Este proceso permite el paso de iones pequeños tales como K+, Na+, Cl-, monosacáridos, aminoácidos y otras moléculas.
Al igual que en la difusión simple el movimiento es a favor del gradiente de concentración de las moléculas. Sin embargo su velocidad de transporte es mayor que el se pronostica con la ley de Fick, ya que no entran en contacto con el centro hidrofóbico de la bicapa. El transporte es específico, transportándose un tipo de moléculas o un grupo de ellas estrechamente relacionados.

La glucosa entra en la mayor parte de las células por difusión facilitada. Parece existir un número limitado de proteínas transportadoras de glucosa. El rápido consumo de la glucosa por la célula (por la tan conocida glicólisis) mantiene el gradiente de concentración. Sin embargo, cuando la concentración externa de glucosa aumenta, la velocidad de transporte no excede cierto límite, sugiriendo una limitación en el transporte.
Transporte activo
Son de interés dos grandes categorías de transporte activo: primario y secundario.
Transporte activo primario
Usa energía (generalmente obtenida de la hidrólisis de ATP), a nivel de la misma proteína de membrana produciendo un cambio conformacional que resulta en el transporte de una molécula a través de la proteína.
El ejemplo mas conocido es la bomba de Na+/K+. La bomba de Na+/K+ realiza un contratransporte (usando un "antyport"). Transporta K+ al interior de la célula y Na+ al exterior de la misma, al mismo tiempo, gastando en el proceso ATP.
En el caso del transporte activo, las proteínas transportadoras deben mover moléculas contra un gradiente de concentración. Por ejemplo en la bomba de sodio-potasio de las células nerviosas el Na+ es mantenido a bajas concentraciones en el interior de las células y el K+ a altas concentraciones (ver cuadro). Las concentraciones están invertidas en el exterior de las células.
Concentraciones iónicas
El valor del potencial eléctrico generado por la diferencia de permeabilidad de los iones y su distribución a ambos lados de la membrana es de -70 mV, resultando el interior de la célula negativo con respecto al exterior. Una idea de la magnitud de este voltaje lo da el hecho que, si consideramos que la membrana tiene un espesor de alrededor de 3,5 nm, el gradiente de voltaje a través de le membrana sería de 0,07 V/ 3,5 x 10 -7cm o sea 200.000 V/cm.
Las bombas iónicas activadas por ATP generan y mantienen gradientes iónicos a través de la membrana plasmática.
Cuando se propaga un mensaje nervioso los iones pasan a través de la membrana (sale el potasio y entra el sodio) transmitiendo el mensaje. Luego de este proceso, los iones deben ser transportados activamente a la "posición de partida" a lo largo de la membrana. Cerca de un tercio del ATP utilizado por un animal en reposo se consume para mantener la bomba Na+/K+.
Existen cuatro tipos de "bombas" que utilizan ATP para mover solutos contra gradientes de concentración.
Las bombas de clase P (compuestas por dos polipéptidos diferentes: alfa y beta) intervienen en el transporte de H+, Na+, K+ y Ca++, se las encuentra en .
las membranas plasmáticas de vegetales, hongos y bacterias , donde
En las membranas plasmáticas de eucariotas actúan como bombas de Na+/ K+ y H+/K+ (células gástricas) y este tipo se lo encuentra además como bombas de Ca++ en dichas células (y en el retículo sarcoplásmico de células musculares)
Las bombas de clase V (múltiples subunidades de membrana y citosólicas) intervienen en el transporte de H+ de membranas de vacuolas de vegetales y hongos, en los endosomas y lisosomas de células animales y en ciertas células animales secretoras de ácidos (p.ej. osteoclastos)
Las bombas de clase ABC (del inglés ATP Binding Cassete, con dos dominios de trasmembrana y dos citosólicos) intervienen en el transporte de iones y moléculas pequeñas. Se encuentran en membranas plasmáticas bacterianas como permeasas asociadas al transporte de aminoácidos y monosacáridos. En retículo endoplásmico de células de mamíferos asociadas al transporte de péptidos relacionados con la presentación de antígenos por las proteínas de CHM, y en membrana plasmática de mamíferos asociadas al transporte de moléculas pequeñas, fosfolípidos y fármacos lipidosímiles pequeños. Esta superfamilia incluye a la MDR1(del inglés MultiDrug Resistence) que exporta un gran número de fármacos y es factor clave de la resistencia de las células cancerosas a los quimioterápicos y a la proteína CFTR (proteína reguladora transmembrana de la fibrosis quística), un canal para el Cl- que es defectuosa en la fibrosis quística.
Transporte activo secundario
Utiliza la energía para establecer un gradiente a través de la membrana celular, y luego utiliza ese gradiente para transportar una molécula de interés contra su gradiente de concentración.
Un ejemplo de ese mecanismo es el siguiente: Escherichia coli establece un gradiente de protones (H+) entre ambos lados de la membrana utilizando energía para bombear protones hacia afuera de la célula. Luego estos protones se acoplan a la lactosa (un azúcar que sirve de nutriente al microorganismo) a nivel de la lactosa-permeasa (otra proteína de transmembrana), la lactosa permeasa usa la energía del protón moviéndose a favor de su gradiente de concentración para transportar la lactosa dentro de la célula.
Este transporte acoplado en la misma dirección a través de la membrana celular se denomina cotransporte (utiliza las proteínas "symport"). Escherichia coli utiliza este tipo de mecanismo para transportar otros azucares tales como ribosa y arabinosa, como así también numerosos aminoácidos.



Transporte mediado por vesículas
1. Fagocitosis: en este proceso, la célula crea una proyecciones de la membrana y el citosol llamadas pseudopodos que rodean la partícula sólida. Una vez rodeada, los pseudopodos se fusionan formando una vesícula alrededor de la partícula llamada vesícula fagocítica o fagosoma. El material sólido dentro de la vesícula es seguidamente digerido por enzimas liberadas por los lisosomas. Los glóbulos blancos constituyen el ejemplo más notable de células que fagocitan bacterias y otras sustancias extrañas como mecanismo de defensa
2. Pinocitosis: en este proceso, la sustancia a transportar es una gotita o vesícula de líquido extracelular. En este caso, no se forman pseudópodos, sino que la membrana se repliega creando una vesícula pinocítica. Una vez que el contenido de la vesícula ha sido procesado, la membrana de la vesícula vuelve a la superficie de la célula.
De esta forma hay un tráfico constante de membranas entre la superficie de la célula y su interior
3. En la endocitosis las partículas hacen que la membrana celular se invagine y luego forme una vesícula que se dirige al interior.


La membrana celular actúa como barrera semipermeable impidiendo la entrada de la mayor parte de las moléculas, dejando pasar selectivamente a otras.
- potencial de agua
- difusión
- ósmosis
- Potencial de agua
La difusión es el movimiento neto de sustancia (líquida o gaseosa) de un área de alta concentración a una de baja concentración. Dado que las moléculas de cualquier sustancia se encuentran en movimiento cuando su temperatura esta por encima de cero absoluto (0 grados Kelvin, o -273 oC), existe una disponibilidad de energía para que las mismas se muevan desde un estado de potencial alto a uno de potencial bajo. La mayoría de las moléculas se mueven desde una concentración alta a una baja, es decir el movimiento neto es desde altas concentraciones a bajas concentraciones.

Eventualmente, si no se agrega energía al sistema las moléculas llegan a un estado de equilibrio en el cual se encuentran distribuidas homogéneamente en el sistema.
Entre las pocas moléculas simples que pueden cruzar la membrana celular por difusión se encuentran gases como nitrógeno, el anhídrido carbónico y el oxígeno. También difunden moléculas sin carga como el etanol y resulta ligeramente permeable al agua y a la urea.

Donde K es la constante de equilibrio de la partición entre aceite y agua. (para el caso de la membrana se analiza Cm, la concentración en el centro de la bicapa hidrófoba).
siendo P proporcional a K

y donde D (característico tanto del soluto como del medio en el que se disuelve) es el coeficiente de difusión de la molécula dentro de la membrana y x el espesor de la misma. Reemplazando nos queda

Por lo tanto la velocidad de difusión es directamente proporcional tanto a la constante de difusión (D) como a la de partición (K) e inversamente proporcional al espesor de la membrana (x). Dado que x prácticamente no varía siendo de alrededor de 3 nm. y, por otra parte las diferencia de los coeficientes de difusión D no suelen ser significativo (para bajas concentraciones se mantiene prácticamente constante). Considerando para un área A constante podríamos simplificar mas y dejar

por lo tanto la velocidad con que difunden las moléculas depende de su coeficiente de partición y de la diferencia de concentración ambos lados de la membrana. En otras palabras cuanto mas hidrófoba sea la molécula, con mayor velocidad difundirá por la membrana plasmática y además esta velocidad se incrementará cuanto mayor sea la diferencia de concentración de las moléculas entre ambos lados de la membrana. Estas relaciones valen para interpretar la difusión de gases y moléculas pequeñas sin carga como el etanol a través de la membrana plasmática.
Por ósmosis se conoce al fenómeno de difusión de agua a través de una membrana semipermeable (conocidas también como de permeabilidad diferencial o de permeabilidad selectiva).

donde p es presión osmótica medida en atmósferas (atm), R la constante de los gases, T la temperatura absoluta y DC la diferencia de las concentraciones de solutos a ambos lados de la membrana.

Las células animales se hinchan cuando son colocadas en soluciones hipotónica, algunas como los eritrocitos terminan estallando debido al agua que penetra en ellas por flujo osmótico (se lisan),
Debido a a su interior hidrofóbico, la bicapa lipídica de una célula constituye una barrera altamente impermeable a la mayoría de las moléculas polares. Esta función de barrera tiene gran importancia ya que le permite a la célula mantener en su citosol a ciertos solutos a concentraciones diferentes a las que están en el fluido extracelular; lo mismo ocurre en cada compartimiento intracelular envuelto por una membrana. El desarrollo evolutivo ha creado sistemas celulares destinados transportar específicamente moléculas hidrosolubles, subsanando el problema del aislamiento celular.

Agre probó su hipótesis en un experimento simple donde él comparó células que tenían la proteína en cuestión con células que no lo tenían. Cuando las células se pusieron en una solución de agua, aquéllas que tenían la proteína en sus membranas absorbieron el agua por ósmosis y se inflaron, mientras aquéllas que carecen de la proteína no eran afectadas en absoluto. Agre también ejecutó los ensayos con las células artificiales, llamadas liposomas ( son un tipo de burbuja de jabón por fuera y el interior constituido por agua). Él encontró que los liposomas se volvieron permeables al agua si la proteína se incrustaba en sus membranas.

bombas: utilizan energía (provista por el ATP) para transportar moléculas contra un gradiente de concentración

transportadores: este tipo de proteínas, luego de fijar las moléculas a transportar (A),


Transporte Activo y Pasivo
Para el transporte pasivo no se requiere que la célula gaste energía. Entre los ejemplos de este tipo de transporte se incluyen la difusión de oxígeno y anhídrido carbónico, la ósmosis del agua y la difusión facilitada.
La difusión facilitada se realiza tanto por medio de las proteínas canal como por los "uniport". Permite que moléculas que de otra manera no podrían atravesar la membrana, difundan libremente hacia afuera y adentro de la célula.

La velocidad de transporte en la difusión facilitado esta limitada por el numero de canales disponibles en la membrana. La velocidad de transporte se satura cuando todos los transportadores están funcionando a su máxima capacidad (ver que la curva indica una "saturación") mientras que en la difusión simple la velocidad de depende solo del gradiente de concentración.
El transporte activo requiere un gasto de energía para transportar la molécula de un lado al otro de la membrana, pero el transporte activo es el único que puede transportar moléculas contra un gradiente de concentración, al igual que la difusión facilitada el transporte activo esta limitado por el numero de proteínas transportadoras presentes.
Ión Interior de la Célula Sangre
Sodio 12 mM 145 mM
Potasio 140 mM 4 mM
La bombas clase F

El mecanismo de transporte secundario Na+-glucosa
Otro sistema de transporte secundario usa la bomba de Sodio/Potasio en una primera etapa, genera así un fuerte gradiente de Sodio a través de la membrana. Luego la proteína "simport" para el sistema Sodio-Glucosa usa la energía del gradiente de Sodio para transportar Glucosa al interior de la célula.

Este sistema se usa de manera original en las células epiteliales del intestino. Estas células toman glucosa y sodio del intestino y lo transportan al torrente sanguíneo utilizando la acción concertada de los "simport" para Sodio/Glucosa, la glucosa permeasa ( una proteína canal de difusión facilitada para la glucosa) y las bombas de Sodio/Potasio.
Se debe hacer notar que las células del intestino se encuentran unidas entre sí por "uniones oclusivas" que impiden que nada proveniente del intestino pase al torrente sanguíneo sin ser primero filtradas por las células epiteliales.

Este mecanismo es la base de la formulación de las "sales de rehidratación oral" que contiene glucosa y electrolitos, utilizada para combatir los efectos de la diarrea provocada por el cólera y otras bacterias patógenas. La cura implica no solamente el uso de antibióticos sino además la reposición del agua que se pierde por acción de las toxinas microbianas. La administración oral de glucosa y sales, que se absorben coordinadamente a nivel del intestino delgado, genera un gradiente osmótico que arrastra el agua desde la luz del intestino hacia la sangre.
Las vesículas y vacuolas que se fusionan con la membrana celular pueden utilizarse para el transporte y liberación de productos químicos hacia el exterior de la célula o para permitir que los mismos entren en la célula. Se aplica el término exocitosis cuando el transporte es hacia fuera de la célula.
Endocitosis: es el proceso mediante el cual la sustancia es transportada al interior de la célula a través de la membrana. Se conocen tres tipos de endocitosis:

4. Endocitosis mediada por receptor: este es un proceso similar a la pinocitosis, con la salvedad que la invaginación de la membrana sólo tiene lugar cuando una determinada molécula, llamada ligando, se une al receptor existente en la membrana (1, 2). Las "fositas recubiertas" ("coated pits") son invaginaciones de la membrana donde se encuentran los receptores.
Captación de la LDL (lipoproteína que contiene ésteres de colesterol)

Una vez formada la vesícula endocítica (3), que al formarse queda recubierta por una proteína llamada clatrina, está se despoja de la clatrina (4) y se une a otras vesículas para formar una estructura mayor llamada endosoma (5). Dentro del endosoma se produce la separación del ligando y del receptor (6): Los receptores son separados y devueltos a la membrana (7, 8), mientras que el ligando se fusiona con un lisosoma siendo digerido por las enzimas de este último (7´, 8´).
Suscribirse a:
Comentarios (Atom)